Received: Fri 05, Jul 2024
Accepted: Fri 19, Jul 2024
Abstract
Background: Different from single-pulse transcranial magnetic stimulation (TMS), repetitive TMS (rTMS) is able to change and modulate cortical activity beyond the stimulation period. This after-effect renders rTMS therapeutic efficacy on neurological and psychiatric disorders.
Methods: We conducted a comprehensive review of clinical studies employing rTMS as a therapeutic approach to treating patients with neuropsychiatric disorders, preclinical studies with animals examining neurobiological changes in brain tissue following exposure to rTMS, and studies with cultured cells exploring cellular and molecular mechanisms underlying TMS-induced changes, in addition to introducing up-to-date knowledge about glia cells.
Results: Findings revealed that rTMS promotes the transformation of microglia from M1 to M2 phenotype and stimulates the astrocyte switch from A1 to A2 phenotype in animal models of neurological and psychiatric diseases. In both in vivo and in vitro experiments, rTMS enhances the adaptive myelination of oligodendrocytes. Along with these changes, rTMS improves behavioral performances in animal models of neuropsychiatric diseases. All of these after-effects last for a relatively long period of time exceeding the exposure time of rTMS.
Conclusion: The after-effects of rTMS on glial cells constitute the neurobiological mechanism for the therapeutic efficacy of this physical intervention in patients with neuropsychiatric disorders.
Keywords
After-effects, glia cells, neuropsychiatric disorders, rTMS, therapeutic efficacy
1. Introduction
Transcranial magnetic stimulation (TMS) is a non-invasive technique based on the principles of electromagnetic induction by which a brief and rapidly changing electric current induces a fluctuating magnetic field that readily penetrates the skull and then induces a second electric current within the excitable tissues of the brain [1]. The induced current may depolarize cortical neuronal assemblies located directly underneath the coil, even in nearby and remote brain regions, thereby exerting therapeutic effects [2]. This technique was introduced to clinical practice in 1985 by Barker and colleagues [3]. Over the past decades, it has become one of the most promising physical therapeutic approaches due to its safety, non-invasion, minimal adverse reactions, effectiveness, and easily-operation [4]. The neuropsychiatric disorders that have been treated by this technique include acute mania, bipolar disorders, catatonia, depression, drug craving, hallucinations, obsessions/compulsions, panic, post-traumatic stress disorder, and schizophrenia, in addition to fibromyalgia, multiple sclerosis, neuropathic pain, Parkinson’s disease, and stroke [4-10].
Applying a single TMS pulse over the primary motor cortex (M1) leads to a twitch in the target muscle along with a motor-evoked potential that can be recorded via surface electromyography [3, 11, 12]. With the same electrophysiological mechanism, applying a TMS pulse to the primary visual cortex can induce a brief flash of light, called phosphene, in the location of the visual field [13-16]. It is currently believed that the direct effect of TMS is to change the membrane potential of target neurons. The indirect effects of TMS, however, may involve more biological mechanisms, including neurotransmitter release, synaptic plasticity, cell survival, as well as neuroinflammatory and immune processes [17].
Different from single-pulse TMS, repetitive TMS (rTMS) is able to change and modulate cortical activity beyond the stimulation period, which is called after-effects. rTMS protocols can be further categorized into single stimulus repeated at fixed inter-stimulus intervals (ISIs, fixed frequency) and patterned protocols that use a combination of different ISIs [18]. Among patterned rTMS protocols, theta-burst stimulation (TBS) (bursts of three pulses at 50-Hz repeated at 200-ms intervals) induces longer-lasting after-effects with shorter application time than conventional rTMS paradigms [19, 20]. It is believed that rTMS exerts its therapeutic effect on neurological and psychiatric disorders through the after-effects including gene activation/regulation, de novo protein expression, morphological changes, changes in intrinsic firing properties and modified network properties resulting from changed inhibition, homeostatic processes and glial functions [21, 22].
This review article focuses on the after-effects of rTMS on glial cells in the brain in order to provide a latest version of the neurobiological mechanisms underlying long-term after-effects of rTMS on brain functions.
2. rTMS Promotes the Transformation of Microglia from M1 to M2 Phenotype
Microglia are the resident immune cells in the brain. They monitor the brain microenvironment by sensing and regulating neuronal activity [22-24], and maintain the brain homeostasis by removing the necrotic cells and debris timely [25]. Microglial activation as defined by the number and morphological changes has been considered to play important roles in pathologic brain states such as various neuropsychiatric disorders [26, 27].
By combining in vivo localized proton magnetic resonance spectroscopy (1H-MRS) and postmortem histological analysis, an early study reported that the application of high intensity, low frequency rTMS does not affect microglial number in the motor cortex or hippocampus in normal healthy rats [28], demonstrating the safety of this physical intervention. Later on, an extremely low-frequency magnetic field (ELF-MF) showed neuroprotective function in an animal model of global cerebral ischemia by decreasing neuronal death and inhibiting activation of astrocytes and microglia [29]. In another animal study, high intensity and frequency rTMS attenuated microglial activation in the injured spinal cord [30].
In a recent animal study, intermittent TBS (iTBS) improved motor function by regulating microglial polarization via toll-like receptor 4 (TLR4)/nuclear factor κ-B (NF-κB)/NOD-like receptor thermal protein domain associated protein 3 (NLRP3) signaling pathway in cerebral ischemic mice [31]. Moreover, rTMS exerted robust neuroprotection in a rat stroke model via altering inflammatory and oxidative status and preserving mitochondrial integrity in the peri-infarct zone [32]. These effects of rTMS on microglia and neuroinflammation in preclinical studies may account for clinical benefits of rTMS for stroke rehabilitation in humans [6]. Indeed, high frequency rTMS applied on the affected dorsolateral prefrontal cortex (DLPFC) of stroke patients was shown to significantly reduce the mRNA level of pro-inflammatory cytokines, including interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α, in blood samples of the patients [33].
Neuroinflammation involves in the pathogenesis of psychiatric disorders such as major depressive disorder, schizophrenia and psychotic disorders, substance use, as well as obsessive-compulsive disorder [34, 35]. Therefore, the therapeutic effects of rTMS on clinical manifestations of patients with psychiatric disorders may be related to the anti-inflammation action of it as demonstrated in the aforementioned animal studies. Of the few preclinical studies exploring therapeutic effects of rTMS in animal models of psychiatric disorders, the following ones provide supporting evidence for the above inference. Tian et al. reported that rTMS improves anxiety and depression-like behaviors and reduces levels of inflammatory factors TNF-α, inducible nitric oxide synthase (iNOS), IL-1β, and IL-6 in the hippocampus of rats subjected to chronic unpredictable mild stress (CUMS) [36].
In another animal study, rTMS (15 Hz) ameliorated depression-like behaviors in CUMS mice while reversed the CUMS-induced neuroinflammation by promoting the polarization transformation of microglia from pro-inflammatory M1 to the anti-inflammatory M2 phenotype in the mice [37]. Moreover, deep rTMS effectively reduced behavioral anomalies while blocking myelin breakdown and oligodendrocyte (OL) loss in cuprizone-fed mice, along with dampened microglia activation and rectified cytokine levels (IL-1β, IL-6, and IL-10) at lesion sites [38]. The cuprizone-fed mouse has been accepted as an animal model of schizophrenia for its relevance to the white matter abnormality and neuroinflammation in the brain of patients with schizophrenia [39-41]. Taken together, these data from animal studies provide new insights into the mechanism for the therapeutic effects of rTMS on clinical manifestations of patients with psychiatric disorders, some of which involve neuroinflammation with microglia as a critical player (Figure 1) [34].
Consistent with the foregoing preclinical studies, rTMS applied to the left DLPFC at 10 Hz was shown to reduce the serum levels of IL-1β and TNF-α in elderly patients with refractory depression in a human study. And this change correlated with Hamilton depression rating scale scores [42]. In a pilot study with a small sample size, the partial changes in cognitive function following rTMS treatment were significantly correlated with IL-1β level changes, although the treatment did not significantly change the serum levels of proinflammatory cytokines (IL-1β, IL-6 and TNF-α) in treatment-resistant depression [43]. Notably, another human study did not detect any significant effects of TBS on the cytokine panels or any correlations between improvement in depressive symptoms and changes in serum inflammatory markers [44]. These results are not consistent with previous studies reporting decreased serum levels of inflammatory cytokines after antidepressant [45-47] or TMS treatment [42]. In another random, double blind, sham controlled study, rTMS treatment for 4 weeks caused a significant increase of IL-2 and C-reactive protein (CRP) levels, while sham-rTMS resulted in an increase of interferon-γ (IFN-γ) levels [48]. These inconsistent results from human studies indicate the complexity of the pathogenesis of human mental disorders and call for further studies to explore the underlying mechanisms for the seemingly conflicts.
3. rTMS Stimulates the Astrocyte Switch from A1 to A2 Phenotype
Astrocytes are the most abundant glial cells in the brain and serve a basic role in the maintenance of the neuronal microenvironment and homeostasis. In addition to providing metabolic and physical supports to neurons, astrocytes are the primary source of energy for neurons and involve in neuronal and synaptic signaling [49, 50]. They are able to respond to neuronal activity with calcium motivation and release of gliotransmitters which further impact synaptic activity [51] thus play important roles in the pathogenesis of neuropsychiatric disorders.
In an early in vitro study, the acute magnetic stimulation (10 Hz for 10 s) given to cultured astrocytes induced a transient increase in glial fibrillary acidic protein (GFAP) level that lasts for 3 days [52]. In a recent study, 1 Hz magnetic stimulation resulted in a significant rise in intracellular calcium in the cytoplasmic and nuclear compartments of the cultured astrocytes [53]. The same group also reported that low intensity repetitive magnetic stimulation reduces expression of genes related to inflammation and calcium signaling in cultured mouse cortical astrocytes [54]. These findings demonstrate the ability of 1 Hz repetitive magnetic stimulation to modulate specific aspects of astrocytic phenotype, potentially contributing to the known effects of low intensity rTMS on excitability and neuroplasticity of brain cells.
Consistent with the above interpretation, rTMS at an extremely low-frequency magnetic field showed neuroprotective functions in the hippocampus in a model of global cerebral ischemia through reduction of neuronal death and inhibiting activation of astrocytes and microglial cells [29]. Similarly, rTMS attenuated astrocyte activation at the site of spinal cord injury in rats [30]. In an animal model of the experimental autoimmune encephalomyelitis (EAE), TMS modified astrogliosis, cell density and lipopolysaccharide levels [55]. In an animal model of neuropathic pain, high-frequency rTMS relieved neuropathic pain through down-regulating the over-expression of nNOS in ipsilateral dorsal root ganglia (DRG) and inhibiting the activity and proliferation of astrocytes in L4-6 spinal dorsal horn ipsilateral to the neuropathic pain [56].
Moreover, high frequency rTMS improved functional recovery of ischemic rats by inhibiting neurotoxic polarization of astrocytes [57]. And rTMS was shown to suppress the excessive astrocyte-vessel interactions while promoting the astrocyte switch from A1 to A2 in the peri-infarct cortex after stroke (Figure 2) [32]. In a more recent animal study, high frequency rTMS improved depression-like behavior, attenuated neural apoptosis, and reversed reduction of astrocytes in a rat model of CUMS [58]. Importantly, high-frequency rTMS for 3 days applied in healthy rats did not alter astroglia reactivity or inflammatory responses such as microglia proliferation while the same treatment up-regulated neuronal metabolic activity in limbic brain structures, along with higher c-Fos levels in the nearest cortical area, providing further evidence for the effectiveness and safety of rTMS as a brain modulation therapy [59].
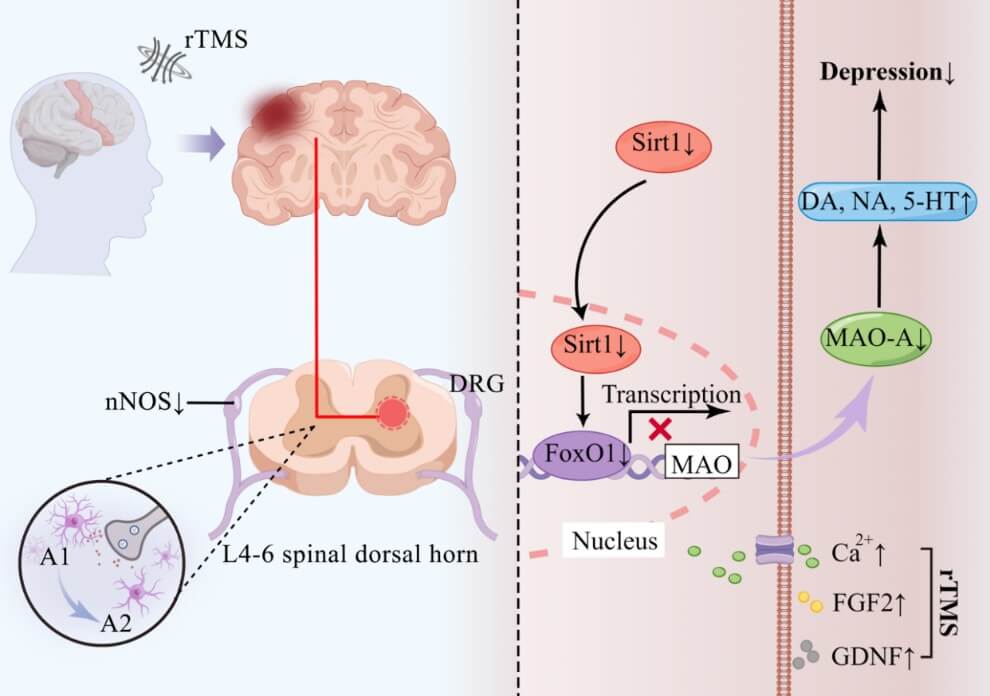
Intriguingly, high frequency (5/10 Hz) rTMS for 7 days improved CUMS-induced depressive-like behaviors, increased 5-hydroxytryptamine (5-HT), dopamine (DA) and norepinephrine (NE) levels, decreased the 5-hydroxyindole acetic acid (5-HIAA) level, lowered sirtuin1 (Sirt1) and monoamine oxidase-A (MAO-A) expression, and reduced MAO-A activity in the PFC [60]. Moreover, rTMS ameliorated depression/anxiety-like behaviors in an animal model of EAE by inhibiting neurotoxic reactive astrocytes in a recent animal study [61]. In a human study, high frequency rTMS applied to the left DLPFC potentiated glutamatergic neurotransmission in depressed adolescents [62]. In patients with PD, high frequency rTMS regulated astrocyte activation by modulating the endocannabinoid system [63]. Taken together, these data provide evidence for the involvement of astrocytes in mediating effects of TMS on synaptic structure and transmission (Figure 2).
4. rTMS Enhances the Adaptive Myelination of Oligodendrocytes
OLs are the neural cells myelinating axons in the central nervous system (CNS). They arise from oligodendrocyte precursor cells (OPC) and follow a specific sequential process to become fully differentiated and functional. These glial cells play important roles in neuroplasticity, neuron nourishment, myelination and nerve repair, in addition to helping maintain normal physiological and adaptive functions of the brain [64, 65]. There are increasing number of reports on OL dysfunction associated with major psychiatric diseases such as schizophrenia [66].
Adaptive myelination, defined as experience-driven oligodendrogenesis and myelination, has been suggested as a significant and lifelong form of neural plasticity which can impact various CNS functions by building a rapport between neurons and oligodendroglia [67]. Accumulating evidence indicates that rTMS has the potential to regulate the process of adaptive myelination. For instance, low-field magnetic stimulation (LFMS, a novel transcranial neuromodulation technology that has shown promising therapeutic potential for a variety of neuropsychiatric conditions) promoted the differentiation of OPCs via non-canonical TGF-β signaling pathways [68]. Relevantly, another in vitro study reported that low frequency pulsed electromagnetic field (PEMF) promoted the differentiation of OPCs, up-regulated the expression level of miR-219-5p and down-regulated Lingo1 during the differentiation of OPCs [69].
Furthermore, in vivo studies found that low-intensity transcranial magnetic stimulation [LI-TMS, delivered as 600 pulses of 10 Hz (60 s), iTBS (192 s), or cTBS (40 s)] promoted the survival and maturation of newborn OLs in the adult mouse brain [70]. In a more recent study, 5 Hz rTMS treatment improved depression-like behavior, attenuated neural apoptosis, and increased the number of Olig2-positive cells in the hilus of the dentate gyrus and the prefrontal cortex in a rat model of depression [71]. All these studies demonstrate that rTMS possesses the potential to influence the adaptive myelination process of OLs, although it is rTMS parameter- and context-dependent.
5. rTMS Effects on OLs in Indirect and Direct Ways
The rTMS inherently carries an associated electric field that can induce electrical and chemical changes in excitable cells such as neurons. Given the tight contact between neuronal axons and membranous processes of OLs, the effects of rTMS on neurons may be subsequently transmitted to OLs exemplified by a pioneer study showing that synaptic inputs could induce cellular depolarization in OPCs and affect the calcium signaling cascades [72]. A later study reported that daily 1 h sessions of electrical stimulation (ES) promoted myelin segment formation in mixed cultures of DRG neurons and OLs, and exposure of DRG neurons to ES prior to OL addition enhanced myelin segment formation, suggesting that ES promotes myelination in a manner involving neuron-specific signaling [73].
In animal studies, neural stimulation in the mouse brain enhanced OPC proliferation, differentiation and myelination within the underlying white matter [74]. Moreover, optogenetic stimulation to neurons in the pre-motor cortex of mammalian brain promoted OPC proliferation, differentiation and thicker myelin formation while increased neural activity [75]. Likewise, in vivo optogenetic stimulation to neurons was shown to induce OPC proliferation, differentiation and remyelination in demyelinated lesions [76]. As for mature OLs, they can sense and respond to neuronal activity, as well as adjust the properties of their myelin sheath to modulate conduction velocity [77, 78]. Taken together, the aforementioned previous studies demonstrate that a stimulation to neurons may indirectly act on the local OLs following the rTMS-induced changes in neuronal activity.
Further evidence supporting this conclusion come as the follows: i) theta burst firing of the pre-synaptic neuron was shown to trigger the insertion of AMPA (calcium-permeable α-amino-3-hydroxy-5-methyl-4-isoxazole-propionicaci) receptors at the OPC postsynaptic density in a process termed glial long-term potentiation (LTP) [79], ii) blocking neuronal action potentials in vivo decreased OPC proliferation [80] and OL production [72], iii) repeated electric stimulation to neurons in the motor cortex promoted the activity-dependent proliferation of OPCs in the corresponding corticospinal tract [81], iv) neuronal activity biases axon selection for myelination in vivo [82], v) the direct current stimulation to neuron-OL co-cultures enhanced the survival and myelinating capacity of the Ols [83], vi) the axonal electrical activity is known to increase OPCs maturation and myelination through axon-derived neurotransmitters: ATP, glutamate, adenosine and GABA (Figure 3) [84].
In addition to the foregoing indirect effects, the direct effects of rTMS on OLs can not be ignored. In an in vitro study by Dolgova et al., a 40-Hz low-field magnetic stimulation (LFMS; 20 min daily for five consecutive days) transiently elevated the level of TGF-β1 in the culture media of CG4 cells, a bipotential glial progenitor cell line that can differentiate into either OL or type 2-astrocyte. In correlation with the TGF-β1 levels, the percentage of cells possessing complex branches and expressing the immature OL marker O4 increased, indicating the accelerated differentiation of CG4 cells towards OLs in the LFMS-exposed cultures [68]. Relevantly, another in vitro study reported that low frequency PEMF promoted the differentiation of OPCs, up-regulated miR-219-5p level and down-regulated the Lingo1 level during the differentiation of OPCs (Figure 3) [69].
6. Molecular Mechanisms Involved in the After-Effects of rTMS on Glial Cells
rTMS can exert effects on glial cells directly or indirectly as demonstrated in the in vitro experiments reviewed above. However, in animals and humans, rTMS definitely functions in both direct and indirect ways involving multiple molecules, as described below.
Accumulating evidence shows that rTMS, especially high frequency rTMS, affects the polarization of microglia and promotes the transformation of microglia into the anti-inflammatory M2 phenotype [31, 32, 36, 85-87]. In vivo and in vitro experimental studies have demonstrated the key roles of NF-κB signaling pathway for rTMS to inhibit the inflammatory phenotype of microglia and promote the anti-inflammatory phenotype. In a recent animal study, early intervention of 20 Hz rTMS suppressed neuroinflammatory responses by regulating the PI3K/Akt/NF-κB signaling pathway in the early stage of 5xFAD mice to decrease pro-inflammation cytokines, thereby alleviated the AD pathology [85]. Similarly, long-term rTMS treatment significantly alleviated inflammatory response, which was accompanied by a decrease in the levels of phosphorylated NF-κB and signal transducer and activator of transcription 6 (STAT6) in a stroke animal model [86].
In another animal study, iTBS was found to promote the transformation of microglia to M2 phenotype and down-regulate the levels of TLR4, NF-κB and p-NF-κB in cerebral ischemic mice, suggesting that iTBS may regulate the polarization of microglia through NF-κB signaling pathway [31]. Moreover, the inhibition of NF-κB signaling pathway in microglia was related to the regulation of let-7b-5p miRNA by rTMS [87]. Taken together, the existing evidence suggests that the inhibition of NF-κB activation may be the final common pathway for rTMS to regulate inflammatory response of microglia under various conditions. In addition, the nuclear factor-E2-related factor 2 (Nrf2) also participates in the rTMS mediated anti-inflammatory effect in a CUMS model [36]. And BDNF-mediated biological response is involved in the regulatory effect of rTMS on microglia [88]. All these data point to a broad spectrum of molecules that may mediate the effect of rTMS on microglia.
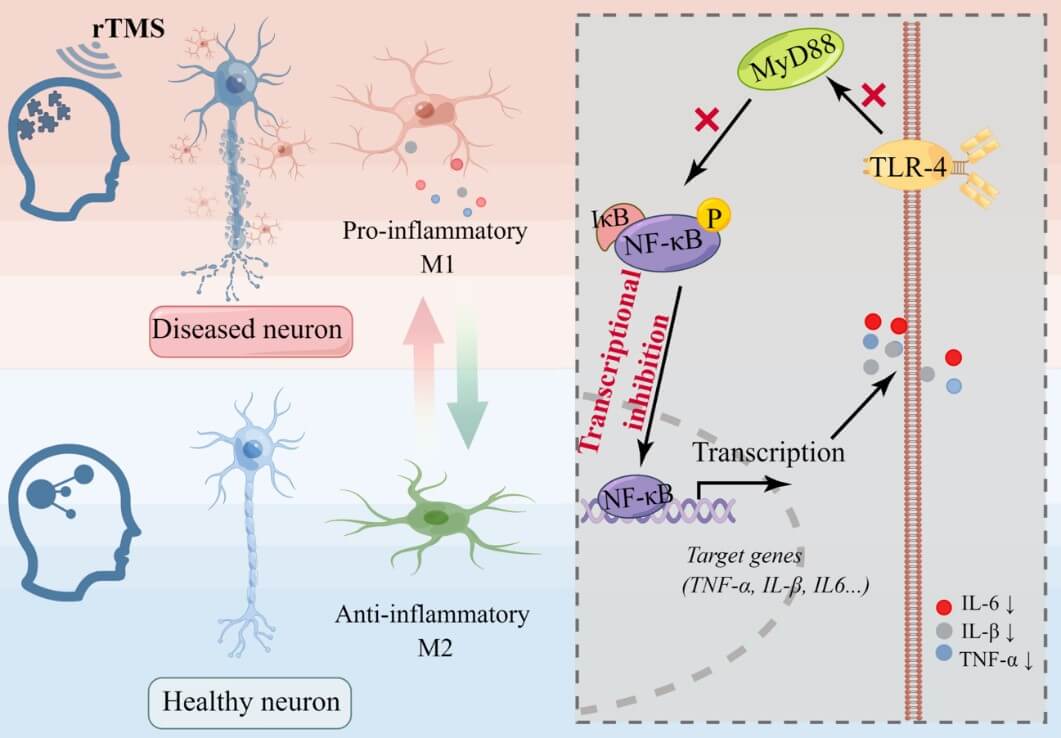
Similar to microglia, astrocyte exists in two types of polarized states of neurotoxic A1 type and neuroprotective A2 type. Recent studies have reported that rTMS promotes the transformation of astrocytes from A1 phenotype to A2 phenotype and reverses the astrogliosis response [32, 57, 61, 63, 89]. Specifically, high frequency rTMS reduced the levels of endocannabinoid receptor 2 (CB2R) in activated astrocytes and its ligands 2-arachidonoylglycerol (2-AG) and anandamide (AEA) in an animal model of PD, suggesting an important role of the endogenous cannabinoid system in the rTMS-induced phenotype transformation of astrocytes [63]. In an in vitro study, rTMS was shown to promote the synthesis and secretion of glial-derived neurotrophic factor (GDNF) by astrocytes [89]. In a CUMS animal model, chronic stress caused a significant increase of apoptosis of astrocyte, which were effectively reversed by rTMS. This reversal effect was accompanied by an increased expression of fibroblast growth factor 2 (FGF2), which mainly comes from astrocytes and promotes the proliferation of astrocyte [58]. Moreover, high frequency rTMS reduced the expression of Sirt1 in astrocytes, thereby regulating the level of monoamine transmitters and exerting antidepressant effect [60]. These previous studies provide experimental evidence for the molecular mechanisms underlying the effects of rTMS on astrocytes dependent on experimental conditions.
Effects of rTMS on OLs and myelination may be achieved in direct, indirect, or both ways as reviewed before. The molecular mechanisms underlying the indirect effects involve some contact-mediated signaling factors and some soluble signaling factors. For instance, Chen et al. reported that neural stimulation increases the axonal location of N-cadherin, a calcium-dependent cell adhesion molecule that may promote myelination as the inhibition of N-cadherin function decreases myelination [90]. Dolgova et al. reported that gamma frequency low-field magnetic stimulation elevates the level of TGF-β1 secretion and accelerates differentiation of OLs in an OL culture model. Moreover, Akt and Erk1/2 signaling proteins involve in the TGF-β-mediated differentiation effect of TMS [91].
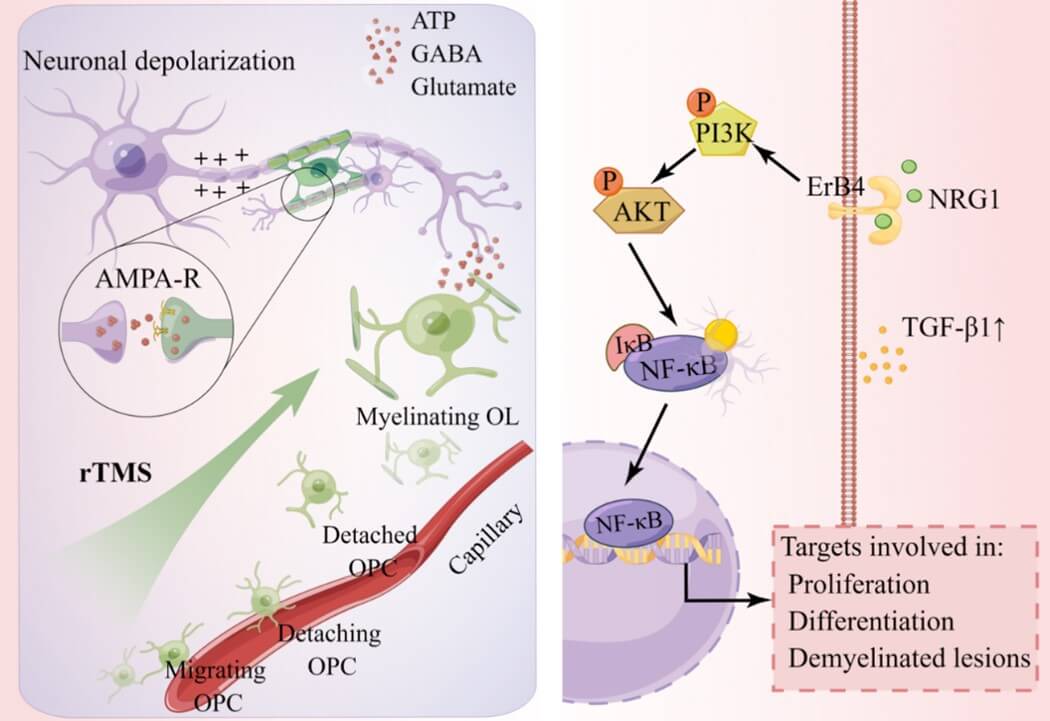
Sun et al. showed that both high frequency and low frequency low field magnetic stimulation repairs the myelin sheath and increase the expression of neuregulin-1 (NRG1) and its receptor ErbB4 in the cuprizone-induced demyelinating mouse model [92]. Both NRG1 and ErbB4 are well-known signaling molecules involved in OL development. Last but not least, rTMS showed a modulatory effect on the glutamate and gamma-aminobutyric acid systems as the treatment increased glutamate levels in the hippocampus and the striatum, but decreased the glutamate concentration in the hypothalamus of rats. The similar changes also happened in gamma-aminobutyric acid concentrations in the same brain regions [93]. Through this effect, rTMS has the potential to facilitate myelination and oligodendrogenesis as glutamate receptors-mediated signaling identified in OLs and OPCs plays an important role in the regulation of activity-dependent myelination [94].
In conclusion, glial cells (microglia, astrocyte, OL) may be affected electrically (directly) and/or chemically (indirectly) by rTMS. Consequently, the normalized glial cells exert their protective effects on brain neurons. It is these after-effects that make rTMS effective in treating neurological and psychiatric disorders. Involving in the neuron-glia interactions are many molecules, exemplified by NF-κB, BDNF and Nrf2 in regulating microglia, CB2R, GDNF and FGF2 in modulating astrocytes, and N-cadherin, TGF-β and NGR1-ErbB4 effecting on OLs (Figures 1-3).
Author Contributions
Haiyun Xu conceived, drafted, and edited this manuscript. Handi Zhang drafted and revised the manuscript. Yutong Li designed and made the figures in the manuscript. All authors approved the final version of the manuscript.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (81971256), Guangdong Province Science and Technology Special Fund (190823085553997), and Li Kashing Foundation (43209502, 2020LKSFG02B). We expressed our gratitude to the above foundations and the support from the Figdraw (Link).
Conflicts of Interest
None.
Data Availability Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
REFERENCES
[1] A T Barker, C W Garnham, I L Freeston
“Magnetic nerve stimulation: the effect of waveform on efficiency,
determination of neural membrane time constants and the measurement of
stimulator output.” Electroencephalogr Clin Neurophysiol Suppl, vol.
43, pp. 227-237, 1991. View at: PubMed
[2] Mark Hallett “Transcranial magnetic
stimulation: a primer.” Neuron, vol. 55, no. 2, pp. 187-199, 2007. View
at: Publisher
Site | PubMed
[3] A T Barker, R Jalinous, I L Freeston
“Non-invasive magnetic stimulation of human motor cortex.” Lancet, vol.
1, no. 8437, pp. 1106-1107, 1985. View at: Publisher Site | PubMed
[4] Simone Rossi, Mark Hallett, Paolo M
Rossini, et al. “Safety, ethical considerations, and application guidelines for
the use of transcranial magnetic stimulation in clinical practice and
research.” Clin Neurophysiol, vol. 120, no. 12, pp. 2008-2039, 2009.
View at: Publisher
Site | PubMed
[5] Stella Rosson, Renato de Filippis,
Giovanni Croatto, et al. “Brain stimulation and other biological
non-pharmacological interventions in mental disorders: An umbrella review.” Neurosci
Biobehav Rev, vol. 139, pp. 104743, 2022. View at: Publisher Site | PubMed
[6] Rongjun Sheng, Changchun Chen, Huan
Chen, et al. “Repetitive transcranial magnetic stimulation for stroke
rehabilitation: insights into the molecular and cellular mechanisms of
neuroinflammation.” Front Immunol, vol. 14, pp. 1197422, 2023.
View at: Publisher
Site | PubMed
[7] Yi-Wen Bai, Qi-Hao Yang, Pei-Jie
Chen, et al. “Repetitive transcranial magnetic stimulation regulates
neuroinflammation in neuropathic pain.” Front Immunol, vol. 14, pp.
1172293, 2023. View at: Publisher
Site | PubMed
[8] Ferran Cuenca-Martínez, Núria
Sempere-Rubio, Sara Mollà-Casanova, et al. “Effects of repetitive-transcranial
magnetic stimulation (rTMS) in fibromyalgia syndrome: An umbrella and mapping
review.” Brain Sci, vol. 13, no. 7, pp. 1059, 2023. View at: Publisher Site | PubMed
[9] Xiaoliang Zhou, Kailin Li, Si Chen,
et al. “Clinical application of transcranial magnetic stimulation in multiple
sclerosis.” Front Immunol, vol. 13, pp. 902658, 2022. View at: Publisher Site | PubMed
[10]
Yi-Xin
Wei, Liang-Dan Tu, Lin He, et al. “Research hotspots and trends of transcranial
magnetic stimulation in Parkinson's disease: a bibliometric analysis.” Front
Neurosci, vol. 17, pp. 1280180, 2023. View at: Publisher Site | PubMed
[11]
V
E Amassian, R Q Cracco “Human cerebral cortical responses to contralateral
transcranial stimulation.” Neurosurgery, vol. 20, no. 1, pp. 148-155,
1987. View at: PubMed
[12]
V
E Amassian, G J Quirk, M Stewart “A comparison of corticospinal activation by
magnetic coil and electrical stimulation of monkey motor cortex.” Electroencephalogr
Clin Neurophysiol, vol. 77, no. 5, pp. 390-401, 1990. View at: Publisher Site | PubMed
[13]
V
E Amassian, R Q Cracco, P J Maccabee, et al. “Suppression of visual perception
by magnetic coil stimulation of human occipital cortex.” Electroencephalogr
Clin Neurophysiol, vol. 74, no. 6, pp. 458-462, 1989. View at: Publisher Site | PubMed
[14]
E
Fernandez, A Alfaro, J M Tormos, et al. “Mapping of the human visual cortex
using image-guided transcranial magnetic stimulation.” Brain Res Brain Res
Protoc, vol. 10, no. 2, pp. 115-124, 2002. View at: Publisher Site | PubMed
[15]
Seth
Elkin-Frankston, Peter J Fried, Alvaro Pascual-Leone, et al. “A novel approach
for documenting phosphenes induced by transcranial magnetic stimulation.” J
Vis Exp, no. 38, pp. 1762, 2010. View at: Publisher Site | PubMed
[16]
Peter
J Fried, Seth Elkin-Frankston, Richard Jarrett Rushmore, et al.
“Characterization of visual percepts evoked by noninvasive stimulation of the
human posterior parietal cortex.” PLoS One, vol. 6, no. 11, pp. e27204,
2011. View at: Publisher
Site | PubMed
[17]
Francisco
Javier Medina, Isaac Túnez “Mechanisms and pathways underlying the therapeutic
effect of transcranial magnetic stimulation.” Rev Neurosci, vol. 24, no.
5, pp. 507-525, 2013. View at: Publisher Site | PubMed
[18]
Noritoshi
Arai, Shingo Okabe, Toshiaki Furubayashi, et al. “Comparison between short
train, monophasic and biphasic repetitive transcranial magnetic stimulation
(rTMS) of the human motor cortex.” Clin Neurophysiol, vol. 116, no. 3,
pp. 605-613, 2005. View at: Publisher Site | PubMed
[19]
Ying-Zu
Huang, Mark J Edwards, Elisabeth Rounis, et al. “Theta burst stimulation of the
human motor cortex.” Neuron, vol. 45, no. 2, pp. 201-206, 2005. View at:
Publisher Site | PubMed
[20]
A
Suppa, Y-Z Huang, K Funke, et al. “Ten years of theta burst stimulation in
humans: Established knowledge, unknowns and prospects.” Brain Stimul,
vol. 9, no. 3, pp. 323-335, 2016. View at: Publisher Site | PubMed
[21]
Wanalee
Klomjai, Rose Katz, Alexandra Lackmy-Vallée, et al. “Basic principles of
transcranial magnetic stimulation (TMS) and repetitive TMS (rTMS).” Ann Phys
Rehabil Med, vol. 58, no. 4, pp. 208-213, 2015. View at: Publisher Site | PubMed
[22]
Dorothy
P Schafer, Emily K Lehrman, Amanda G Kautzman, et al. “Microglia sculpt
postnatal neural circuits in an activity and complement-dependent manner.” Neuron,
vol. 74, no. 4, pp. 691-705, 2012. View at: Publisher Site | PubMed
[23]
Ana
Badimon, Hayley J Strasburger, Pinar Ayata, et al. “Negative feedback control
of neuronal activity by microglia.” Nature, vol. 586, no. 7829, pp.
417-423, 2020. View at: Publisher
Site | PubMed
[24]
Felix
Christopher Nebeling, Stefanie Poll, Lena Christine Justus, et al. “Microglial
motility is modulated by neuronal activity and correlates with dendritic spine
plasticity in the hippocampus of awake mice.” Elife, vol. 12, pp.
e83176, 2023. View at: Publisher
Site | PubMed
[25]
Qingyun
Li, Ben A Barres “Microglia and macrophages in brain homeostasis and disease.”
Nat Rev Immunol, vol. 18, no. 4, pp. 225-242, 2018. View at: Publisher Site | PubMed
[26]
Marco
Prinz, Steffen Jung, Josef Priller “Microglia biology: One century of evolving
concepts.” Cell, vol. 179, no. 2, pp. 292-311, 2019. View at: Publisher Site | PubMed
[27]
Marco
Prinz, Takahiro Masuda, Michael A Wheeler, et al. “Microglia and central
nervous system-associated macrophages-From origin to disease modulation.” Annu
Rev Immunol, vol. 39, pp. 251-277, 2021. View at: Publisher
Site | PubMed
[28]
David
Liebetanz, Susanne Fauser, Thomas Michaelis, et al. “Safety aspects of chronic
low-frequency transcranial magnetic stimulation based on localized proton
magnetic resonance spectroscopy and histology of the rat brain.” J Psychiatr
Res, vol. 37, no. 4, pp. 277-286. View at: Publisher Site | PubMed
[29]
Snežana
Rauš, Vesna Selaković, Milica Manojlović-Stojanoski, et al. Response of hippocampal neurons and glial cells to
alternating magnetic field in gerbils submitted to global cerebral ischemia. Neurotox
Res, vol. 23, no. 1, pp. 79-91, 2013. View at: Publisher Site | PubMed
[30]
Ji
Young Kim, Gyu-Sik Choi, Yun-Woo Cho, et al. “Attenuation of spinal cord
injury-induced astroglial and microglial activation by repetitive transcranial
magnetic stimulation in rats.” J Korean Med Sci, vol. 28, no. 2, pp.
295-299, 2013. View at: Publisher
Site | PubMed
[31]
Lu
Lu, Meixi Liu, Yunhui Fan, et al. “Intermittent theta-burst stimulation
improves motor function by inhibiting neuronal pyroptosis and regulating
microglial polarization via TLR4/NFκB/NLRP3 signaling pathway in cerebral
ischemic mice.” J
Neuroinflammation, vol. 19, no. 1, pp. 141, 2022. View at: Publisher Site | PubMed
[32]
Xuemei
Zong, Yan Dong, Yuyu Li, et al. “Beneficial effects of theta-burst transcranial
magnetic stimulation on stroke injury via improving neuronal microenvironment
and mitochondrial integrity.” Transl Stroke Res, vol. 11, no. 3, pp.
450-467, 2020. View at: Publisher
Site | PubMed
[33]
Byoungwoo
Cha, Jongwook Kim, Jong Moon Kim, et al. “Therapeutic effect of repetitive
transcranial magnetic stimulation for post-stroke vascular cognitive
impairment: A prospective pilot study.” Front Neurol, vol. 13, pp.
813597, 2022. View at: Publisher
Site | PubMed
[34]
Jeffrey
H Meyer, Simon Cervenka, Min-Jeong Kim, et al. “Neuroinflammation in
psychiatric disorders: PET imaging and promising new targets.” Lancet
Psychiatry, vol. 7, no. 12, pp. 1064-1074, 2020. View at: Publisher Site | PubMed
[35]
Bingqi
Guo, Mengyao Zhang, Wensi Hao, et al. “Neuroinflammation mechanisms of
neuromodulation therapies for anxiety and depression”. Transl Psychiatry,
vol. 13, no. 1, pp. 5, 2023. View at: Publisher Site | PubMed
[36]
Li
Tian, Si-Si Sun, Long-Biao Cui, et al. “Repetitive transcranial magnetic
stimulation elicits antidepressant- and anxiolytic-like effect via nuclear
factor-E2-related factor 2-mediated anti-inflammation mechanism in rats.” Neuroscience,
vol. 429, pp. 119-133, 2020. View at: Publisher
Site | PubMed
[37]
Chengchao
Zuo, Huan Cao, Fang Feng, et al. “Repetitive transcranial magnetic stimulation
exerts anti-inflammatory effects via modulating glial activation in mice with
chronic unpredictable mild stress-induced depression.” Int Immunopharmacol,
vol. 109, pp. 108788, 2022. View at: Publisher Site | PubMed
[38]
Liu
Yang, Yawen Su, Fannv Guo, et al. “Deep rTMS mitigates behavioral and
neuropathologic anomalies in cuprizone-exposed mice through reducing microglial
proinflammatory cytokines.” Front Integr Neurosci, vol. 14, pp. 556839,
2020. View at: Publisher
Site | PubMed
[39]
Haiyun
Xu, Hong-Ju Yang, Yanbo Zhang, et al. “Behavioral and neurobiological changes
in C57BL/6 mice exposed to cuprizone.” Behav Neurosci, vol. 123, no. 2,
pp. 418-429, 2009. View at: Publisher
Site | PubMed
[40]
Nicole
R Herring, Christine Konradi “Myelin, copper, and the cuprizone model of
schizophrenia.” Front Biosci (Schol Ed), vol. 3, no. 1, pp. 23-40, 2011.
View at: Publisher Site | PubMed
[41]
Haiyun
Xu, Xin-Min Li “White matter abnormalities and animal models examining a
putative role of altered white matter in schizophrenia.” Schizophr Res
Treatment, vol. 2011, pp. 826976, 2011. View at: Publisher Site | PubMed
[42]
Xiangxiang
Zhao, Yanpeng Li, Qing Tian, et al. “Repetitive transcranial magnetic
stimulation increases serum brain-derived neurotrophic factor and decreases
interleukin-1β and tumor necrosis factor-α in elderly patients with refractory
depression.” J Int Med Res,
vol. 47, no. 5, pp. 1848-1855, 2019. View at: Publisher Site | PubMed
[43]
Hiroshi
Tateishi, Yoshito Mizoguchi 2, Atsushi Kawaguchi, et al. “Changes in
interleukin-1 beta induced by rTMS are significantly correlated with partial
improvement of cognitive dysfunction in treatment-resistant depression: a pilot
study.” Psychiatry Res, vol. 289, pp. 112995, 2020. View at: Publisher Site | PubMed
[44]
Po-Han
Chou, Ming-Kuei Lu, Chon-Haw Tsai, et al. “Antidepressant efficacy and immune
effects of bilateral theta burst stimulation monotherapy in major depression: A
randomized, double-blind, sham-controlled study.” Brain Behav Immun,
vol. 88, pp. 144-150, 2020. View at: Publisher Site | PubMed
[45]
Yekta
Dowlati, Nathan Herrmann, Walter Swardfager, et al. “A meta-analysis of
cytokines in major depression.” Biol Psychiatry, vol. 67, no. 5, pp.
446-457, 2010. View at: Publisher
Site | PubMed
[46]
Christoph
Kraus, Bashkim Kadriu, Rupert Lanzenberger, et al. “Prognosis and improved
outcomes in major depression: A review.” Focus (Am Psychiatr Publ), vol.
18, no. 2, pp. 220-235, 2020. View at: Publisher Site | PubMed
[47]
R
Strawbridge, D Arnone, A Danese, et al. “Inflammation and clinical response to
treatment in depression: A meta-analysis.” Eur Neuropsychopharmacol,
vol. 25, no. 10, pp. 1532-1543, 2015. View at: Publisher Site | PubMed
[48]
Qiang
Wang, Lingyun Zeng, Wenjuan Hong, et al. “Inflammatory cytokines changed in
patients with depression before and after repetitive transcranial magnetic
stimulation treatment.” Front Psychiatry, vol. 13, pp. 925007, 2022.
View at: Publisher
Site | PubMed
[49]
Andrea
Volterra, Jacopo Meldolesi “Astrocytes, from brain glue to communication
elements: the revolution continues.” Nat Rev Neurosci, vol. 6, no. 8,
pp. 626-640, 2005. View at: Publisher
Site | PubMed
[50]
Nicola
J Allen, Ben A Barres “Signaling between glia and neurons: focus on synaptic
plasticity.” Curr Opin Neurobiol, vol. 15, no. 5, pp. 542-548, 2005.
View at: Publisher
Site | PubMed
[51]
Robert
Zorec, Alfonso Araque, Giorgio Carmignoto, et al. “Astroglial excitability and
gliotransmission: an appraisal of Ca2+ as a signalling route.” ASN Neuro,
vol. 4, no. 2, pp. e00080, 2012. View at: Publisher Site | PubMed
[52]
P
Chan, L F Eng, Y L Lee, et al. “Effects of pulsed magnetic stimulation of GFAP
levels in cultured astrocytes.” J Neurosci Res, vol. 55, no. 2, pp.
238-244, 1999. View at: Publisher
Site | PubMed
[53]
Darren
Clarke, Marissa A Penrose, Tamasin Penstone, et al. “Frequency-specific effects
of repetitive magnetic stimulation on primary astrocyte cultures.” Restor
Neurol Neurosci, vol. 35, no. 6, pp. 557-569, 2017. View at: Publisher Site | PubMed
[54]
Darren
Clarke, Jamie Beros, Kristyn A Bates, et al. “Low intensity repetitive magnetic
stimulation reduces expression of genes related to inflammation and calcium
signalling in cultured mouse cortical astrocytes.” Brain Stimul, vol.
14, no. 1, pp. 183-191, 2021. View at: Publisher Site | PubMed
[55]
Francisco
J Medina-Fernández, Evelio Luque, Macarena Aguilar-Luque, et al. “Transcranial
magnetic stimulation modifies astrocytosis, cell density and lipopolysaccharide
levels in experimental autoimmune encephalomyelitis.” Life Sci, vol.
169, pp. 20-26, 2017. View at: Publisher Site | PubMed
[56]
Lu
Yang, Sai-Hua Wang, Yan Hu, et al. “Effects of repetitive transcranial magnetic
stimulation on astrocytes proliferation and nNOS expression in neuropathic pain
rats.” Curr Med Sci, vol. 38, no. 3, pp. 482-490, 2018. View at: Publisher Site | PubMed
[57]
Ye
Hong, Qian Liu, Mengna Peng, et al. “High-frequency repetitive transcranial
magnetic stimulation improves functional recovery by inhibiting neurotoxic
polarization of astrocytes in ischemic rats.” J Neuroinflammation, vol.
17, no. 1, pp. 150, 2020. View at: Publisher Site | PubMed
[58]
Junni
Yan, Fuping Zhang, Le Niu, et al. “High-frequency repetitive transcranial
magnetic stimulation mitigates depression-like behaviors in CUMS-induced rats
via FGF2/FGFR1/p-ERK signaling pathway.” Brain Res Bull, vol. 183, pp.
94-103, 2022. View at: Publisher
Site | PubMed
[59]
Candela
Zorzo, Sara G Higarza, Marta Méndez, et al. “High frequency repetitive
transcranial magnetic stimulation improves neuronal activity without affecting
astrocytes and microglia density.” Brain Res Bull, vol. 150, pp.
13-20, 2019. View at: Publisher
Site | PubMed
[60]
Zheng-Wu
Peng, Fen Xue, Cui-Hong Zhou, et al. “Repetitive transcranial magnetic
stimulation inhibits Sirt1/MAO-A signaling in the prefrontal cortex in a rat
model of depression and cortex-derived astrocytes.” Mol Cell Biochem,
vol. 442, no. 1-2, pp. 59-72, 2018. View at: Publisher Site | PubMed
[61]
Chao
Yu, Yiwen Ruan, Xiaobo Sun, et al. “rTMS ameliorates depression/anxiety-like
behaviors in experimental autoimmune encephalitis by inhibiting neurotoxic
reactive astrocytes.” J Affect Disord, vol. 331, pp. 352-361, 2023. View
at: Publisher Site | PubMed
[62]
Paul
E Croarkin, Paul A Nakonezny, Christopher A Wall, et al. “Transcranial magnetic
stimulation potentiates glutamatergic neurotransmission in depressed
adolescents.” Psychiatry Res Neuroimaging, vol. 247, pp. 25-33, 2016.
View at: Publisher
Site | PubMed
[63]
Xin
Kang, Bing Zhang, Wanqing Du, et al. “High-frequency repetitive transcranial
magnetic stimulation regulates astrocyte activation by modulating the
endocannabinoid system in Parkinson's Disease.” Mol Neurobiol, vol. 59,
no. 8, pp. 5121-5134, 2022. View at: Publisher Site | PubMed
[64]
Klaus-Armin
Nave, Hauke B Werner “Ensheathment and myelination of axons: Evolution of glial
functions.” Annu Rev Neurosci, vol. 44, pp. 197-219, 2021. View at: Publisher
Site | PubMed
[65]
Dominik
Sakry, Jacqueline Trotter “The role of the NG2 proteoglycan in OPC and CNS
network function.” Brain Res, vol. 1638, no. Pt B, pp. 161-166, 2016.
View at: Publisher
Site | PubMed
[66]
Yang-Wen
Ai, Yang Du, Lei Chen, et al. “Glial cell abnormalities in major psychiatric
diseases: A systematic review of postmortem brain studies.” Mol Neurobiol,
vol. 59, no. 3, pp. 1665-1692, 2023. View at: Publisher Site | PubMed
[67]
Sarah
E Pease-Raissi, Jonah R Chan “Building a (w)rapport between neurons and
oligodendroglia: Reciprocal interactions underlying adaptive myelination.” Neuron,
vol. 109, no. 8, pp. 1258-1273, 2021. View at: Publisher Site | PubMed
[68]
Natalia
Dolgova, Zelan Wei, Brandon Spink, et al. “Low-field magnetic stimulation
accelerates the differentiation of oligodendrocyte precursor cells via
non-canonical TGF-β signaling pathways.” Mol Neurobiol, vol. 58, no. 2, pp. 855-866, 2021.
View at: Publisher
Site | PubMed
[69]
Fei
Yao, Ziyu Li, Li Cheng, et al. “Low frequency pulsed electromagnetic field
promotes differentiation of oligodendrocyte precursor cells through
upregulation of miR-219-5p in vitro.” Life Sci, vol. 223, pp.
185-193, 2019. View at: Publisher
Site | PubMed
[70]
Carlie
L Cullen, Matteo Senesi, Alexander D Tang, et al. “Low-intensity transcranial
magnetic stimulation promotes the survival and maturation of newborn
oligodendrocytes in the adult mouse brain.” Glia, vol. 67, no. 8, pp.
1462-1477, 2019. View at: Publisher
Site | PubMed
[71]
Gerardo
Bernabé Ramírez-Rodríguez, David Meneses San-Juan, Allan Irasek Rico-Becerra,
et al. “Repetitive transcranial magnetic stimulation and fluoxetine reverse
depressive-like behavior but with differential effects on Olig2-positive cells
in chronically stressed mice.” Neuropharmacology, vol. 236, pp. 109567,
2023. View at: Publisher
Site | PubMed
[72]
C
Demerens, B Stankoff, M Logak, et al. “Induction of myelination in the central
nervous system by electrical activity.” Proc Natl Acad Sci USA, vol. 93,
no. 18, pp. 9887-9892, 1996. View at: Publisher Site | PubMed
[73]
Misti
Malone, Devin Gary, In Hong Yang, et al. “Neuronal activity promotes
myelination via a cAMP pathway.” Glia, vol. 61, no. 6, pp. 843-854,
2013. View at: Publisher
Site | PubMed
[74]
Stanislaw
Mitew, Ilan Gobius, Laura R Fenlon, et al. “Pharmacogenetic stimulation of
neuronal activity increases myelination in an axon-specific manner.” Nat
Commun, vol. 9, no. 1, pp. 306, 2018. View at: Publisher Site | PubMed
[75]
Erin
M Gibson, David Purger, Christopher W Mount, et al. “Neuronal activity promotes
oligodendrogenesis and adaptive myelination in the mammalian brain.” Science,
vol. 344, no. 6183, pp. 1252304, 2014. View at: Publisher Site | PubMed
[76]
Fernando
C Ortiz, Chloé Habermacher, Mariana Graciarena, et al. “Neuronal activity in
vivo enhances functional myelin repair.” JCI Insight, vol. 5, no. 9,
pp. e123434, 2019. View at: Publisher
Site | PubMed
[77]
Yoshihiko
Yamazaki, Yasukazu Hozumi, Kenya Kaneko, et al. “Modulatory effects of
oligodendrocytes on the conduction velocity of action potentials along axons in
the alveus of the rat hippocampal CA1 region.” Neuron Glia Biol, vol. 3,
no. 4, pp. 325-334, 2007. View at: Publisher Site | PubMed
[78]
Yoshihiko
Yamazaki, Hiroki Fujiwara, Kenya Kaneko, et al. “Short- and long-term
functional plasticity of white matter induced by oligodendrocyte depolarization
in the hippocampus.” Glia, vol. 62, no. 8, pp. 1299-1312, 2014.
View at: Publisher Site | PubMed
[79]
Marzieh
Zonouzi 1, Massimiliano Renzi, Mark Farrant, et al. “Bidirectional plasticity
of calcium-permeable AMPA receptors in oligodendrocyte lineage cells.” Nat
Neurosci, vol. 14, no. 11, pp. 1430-1438, 2011. View at: Publisher Site | PubMed
[80]
B
A Barres, M C Raff “Proliferation of oligodendrocyte precursor cells depends on
electrical activity in axons.” Nature, vol. 361, no. 6409, pp. 258-260,
1993. View at: Publisher
Site | PubMed
[81]
Qun
Li, Marcel Brus-Ramer, John H Martin, et al. “Electrical stimulation of the
medullary pyramid promotes proliferation and differentiation of oligodendrocyte
progenitor cells in the corticospinal tract of the adult rat.” Neurosci Lett,
vol. 479, no. 2, pp. 128-133, 2010. View at: Publisher Site | PubMed
[82]
Jacob
H Hines, Andrew M Ravanelli, Rani Schwindt, et al. “Neuronal activity biases
axon selection for myelination in vivo.” Nat Neurosci, vol. 18,
no. 5, pp. 683-689, 2015. View at: Publisher
Site | PubMed
[83]
Devin
S Gary, Misti Malone, Paul Capestany, et al. “Electrical stimulation promotes
the survival of oligodendrocytes in mixed cortical cultures.” J Neurosci Res,
vol. 90, no. 1, pp. 72-83, 2012. View at: Publisher Site | PubMed
[84]
R
Douglas Fields “A new mechanism of nervous system plasticity:
activity-dependent myelination.” Nat Rev Neurosci, vol. 16, no. 12, pp.
756-767, 2015. View at: Publisher
Site | PubMed
[85]
Kai
Li, Xin Wang, Yu Jiang, et al. “Early intervention attenuates synaptic
plasticity impairment and neuroinflammation in 5xFAD mice.” J Psychiatr Res,
vol. 136, pp. 204-216, 2021. View at: Publisher Site | PubMed
[86]
Jing
Luo, Yuan Feng, Mingyue Li, et al. “Repetitive transcranial magnetic
stimulation improves neurological function and promotes the anti-inflammatory
polarization of microglia in ischemic rats.” Front Cell Neurosci, vol.
16, pp. 878345, 2022. View at: Publisher Site | PubMed
[87]
Ye
Hong, Jinfeng Lyu, Lin Zhu, et al. “High-frequency repetitive transcranial
magnetic stimulation (rTMS) protects against ischemic stroke by inhibiting M1
microglia polarization through let-7b-5p/HMGA2/NF-κB signaling pathway.” BMC Neurosci, vol. 23,
no. 1, pp. 49, 2022. View at: Publisher Site | PubMed
[88]
Ivana
Stevanovic, Bojana Mancic, Tihomir Ilic, et al. “Theta burst stimulation
influence the expression of BDNF in the spinal cord on the experimental
autoimmune encephalomyelitis.” Folia Neuropathol, vol. 57, no. 2,
pp. 129-145, 2019. View at: Publisher
Site | PubMed
[89]
Genilso
Gava-Junior, Susana A Ferreira, Cláudio Roque, et al. “High-frequency
repetitive magnetic stimulation rescues ischemia-injured neurons through
modulation of glial-derived neurotrophic factor present in the astrocyte's
secretome.” J Neurochem, vol. 164, no. 6, pp. 813-828, 2023. View at: Publisher Site | PubMed
[90]
Min
Chen, Yang Xu, Rongchen Huang, et al. “N-Cadherin is involved in neuronal
activity-dependent regulation of myelinating capacity of zebrafish individual
oligodendrocytes in vivo.” Mol Neurobiol, vol. 54, no. 9, pp.
6917-6930, 2017. View at: Publisher
Site | PubMed
[91]
Fei
Yao, Ziyu Li, Li Cheng, et al. “Low frequency pulsed electromagnetic field
promotes differentiation of oligodendrocyte precursor cells through
upregulation of miR-219-5p in vitro.” Life Sci, vol. 223, pp.
185-193, 2019. View at: Publisher
Site | PubMed
[92]
Zuoli
Sun, Tianhe Jiang, Yan Wu, et al. “Low field magnetic stimulation ameliorates
schizophrenia-like behavior and up-regulates neuregulin-1 expression in a mouse
model of cuprizone-induced demyelination.” Front Psychiatry, vol. 9, pp.
675, 2018. View at: Publisher
Site | PubMed
[93] Li Yue, Huo Xiao-lin, Song Tao “The effects of chronic repetitive transcranial magnetic stimulation on glutamate and gamma-aminobutyric acid in rat brain.” Brain Res, vol. 1260, pp. 94-99, 2009. View at: Publisher Site | PubMed
[94] Elliot H Choi, Agata Blasiak, Joonho Lee, et al. “Modulation of neural activity for myelination in the central nervous system.” Front Neurosci, vol. 13, pp. 952, 2019. View at: Publisher Site | PubMed